
Abstract
For decades, mainstream biology has operated under a prevailing assumption: that the body is essentially a machine of biochemical transactions, all directed toward anatomical outcomes. At the heart of this view sits the genome — thousands of genes arranged into DNA — which was believed to govern these reactions by serving as a master blueprint.
This fixation on gene theory gained momentum largely from breakthroughs in sequencing technology, which successfully traced the origins of monogenic diseases — inherited conditions caused by a single defective gene, such as inborn metabolic errors or mutations. Encouraged by these findings, proponents of gene theory made a sweeping leap: they applied the same genomic logic to chronic, degenerative diseases and cancer. For years, the prevailing clinical strategy became identifying faulty genes or mutations and targeting them with therapies or drugs. What began as a framework for rare inherited conditions gradually colonised all of biomedicine, fuelling a push toward mass genomics as the path to better human health.
A natural question emerged: What if the fact that the genome holds the answer to chronic diseases is a flawed approach?
This hypothesis — that sequencing the genome and mapping defective genes would unlock answers to virtually all disease — drove one of science’s most ambitious undertakings: the Human Genome Project, completed in 2003. Yet more than two decades later, the results speak for themselves. The incidence of chronic disease and cancer in India has quadrupled, while over 90% of the US population now shows signs of metabolic dysfunction.
The evidence is hard to ignore: genes simply do not hold the answers to most chronic diseases. And yet, a wave of healthtech startups in India continues to burn investor capital sequencing static genes and mutations, pairing findings with rigid nutritional protocols & pharmaceutical interventions in the hope of correcting what they label genetic faults. The outcomes tell a different story — measurable improvements in patient health remain elusive, with the rare exceptions amounting to little more than anecdotal evidence affecting perhaps 5% of those tested.
A critical distinction is worth emphasising here. The genome is an instruction manual — nothing more. It does not act on its own. It is the cell that reads those instructions and executes biochemical functions, activating the right enzymes and proteins at the right time. This shifts attention toward something far more fundamental: biology runs on energy. Every cellular process demands ATP, and it is the mitochondria — residing in virtually every cell in the body, save for red blood cells — that supply it. Far from being passive powerhouses, mitochondria are increasingly understood as the true command centre of cellular life, orchestrating homeostasis from within.
The research backing this view is substantial. Work by Martin Picard and Doug Wallace demonstrates that mitochondria regulate expression of up to two-thirds of the nuclear genes encoded in the cell’s nucleus.
Meanwhile, Professor Michael Levin’s extensive studies in developmental biology reveal an even more startling picture: cells do not simply execute genetic instructions — they form dynamic, collective networks that sense the external environment, communicate with one another, draw on patterned bioelectric memory, and can actively override or disregard genetic directives when building and repairing tissue. This discovery carries profound implications, particularly as a potential therapeutic frontier in cancer treatment.
This blog explores the emerging innovations in mitochondrial and developmental biology that may catalyse a genuine paradigm shift — one that moves medicine away from its fixation on the hardwired genome and toward reprogramming software- collective intelligence of cells and the central role of mitochondrial biology in addressing our growing epidemic of chronic diseases.
Genome as hardware, not architect
The genome functions as the biochemical hardware of life — encoding proteins and issuing molecular instructions from the top down. But DNA alone cannot specify anatomical form. It does not dictate where your hands, eyes, or head should develop. The genome supplies the tools; it does not execute the plan.
The human body contains roughly 37 trillion cells, each operating as a remarkably capable, semi-autonomous unit. Within their individual cognitive and computational limits, cells make local decisions — yet no single cell inherently knows which organ it belongs to, or where specific structures should appear.
Rather than passive machines executing hardcoded genetic programs, cells are active problem-solvers. They draw on physiological networks, localized memory, and bioelectric signaling to carry out large-scale, goal-directed behavior. Cellular memory extends well beyond genetic code — encompassing epigenetic imprinting, chromatin remodeling, and stored bioelectric states. Together, these define the baseline parameters that guide how cells differentiate and migrate.
Instead of just consulting pre-written genetic blueprints alone, cells form and utilize bioelectric networks to read their physical position, communicate with neighbors, and self-organize into complex tissues.
The Emergence of Collective Intelligence
Biological cells, then, do not simply follow a rigid genetic script. They operate as a collaborative network — continuously reading their environment, exchanging signals, and drawing on shared bioelectric memory to build and repair tissue.
What emerges from this collaboration is something far more powerful: the collective intelligence of a distributed cellular network. When connected through ion channels and gap junctions, cells dramatically expand their shared computational capacity, enabling them to pursue large-scale anatomical goals — with profound implications for fields such as cancer research.
This “cell collective” operates through emergent, system-level intelligence that prioritizes the anatomical coherence of the whole organism over isolated genetic commands. When genetic mutations introduce structural errors, the collective can functionally override them, redirecting developmental processes toward a target morphology.
The Cognitive capacity of cells(1)
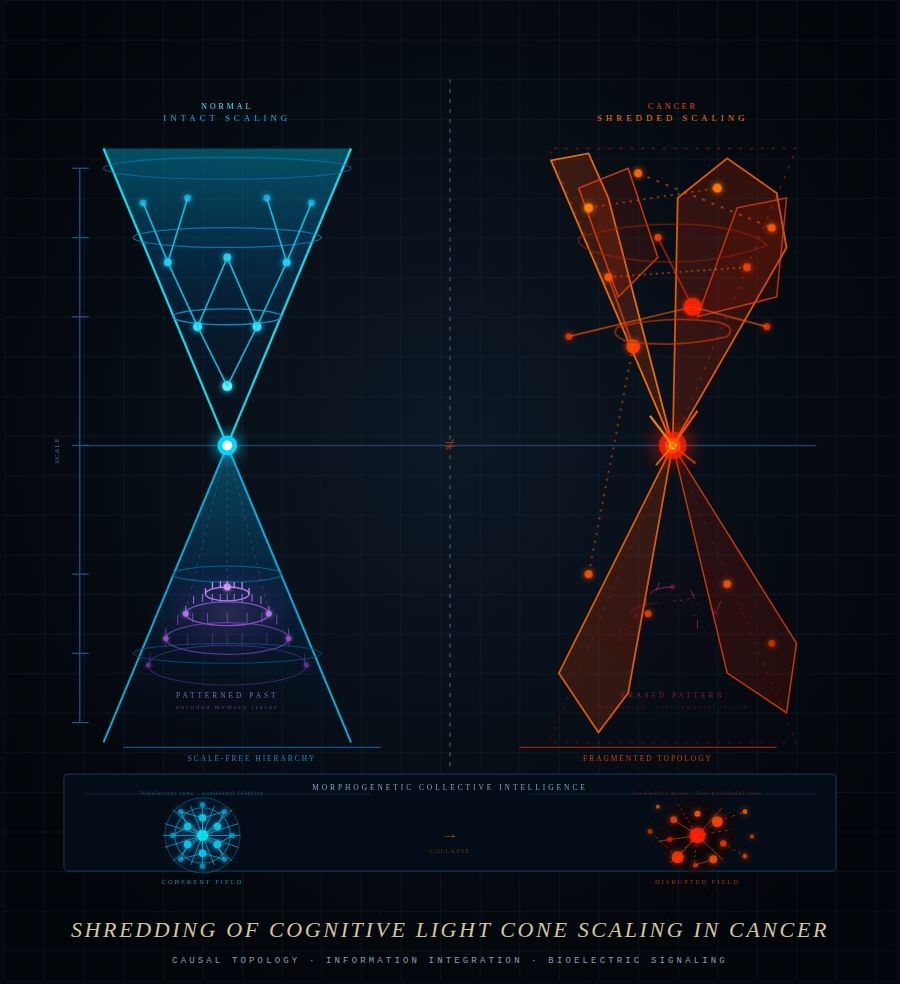
This framework, extensively developed by Professor Michael Levin and supported by a growing body of research, centers on the concept of the cognitive light cone. At the individual level, a cell’s cognitive light cone is narrow — it holds short-term memory, limited predictive capacity, and pursues modest goals such as maintaining local pH. But when cells operate coherently as a bioelectric network, this light cone expands dramatically. The collective gains the ability to pursue goals across far greater scales of space and time than any single cell could achieve alone.
This also illuminates how cells draw on stored memory and DNA-encoded instructions to execute the intricate, coordinated processes that drive morphogenesis — while the collective retains the capacity to override genetic instructions in service of large-scale developmental goals.
Rethinking Disease from collective intelligence perspective
The implications of this paradigm shift are significant. If disease arises not merely from genetic mutation but from a breakdown in cellular communication and collective decision-making, then restoring health means restoring information flow.
Cancer, viewed through this lens, is reframed not simply as rogue genetic replication, but as cells that have dropped out of the collective — reverting to a primitive, individualistic mode of behavior. This opens therapeutic pathways that target the bioelectric and cognitive architecture of tissue itself, potentially offering new strategies for a wide range of chronic and degenerative conditions where gene-centric approaches have reached their limits.
The cell’s adaptive capacity as agent- implications of game theory
Current biological progress has been made at understanding the human body at plasma & to some extent at molecular level ( very few companies in India focus on molecules). The current practice of relying on the genome is poor at understanding & rebuilding the anatomical structure & rather focus has been on identifying faulty genes or protein to fuel drug development that could target these specific molecules in isolation. This is the reason the focus is more on suppressing the adverse biochemical reactions while not developing regenerative therapeutics or signalling molecules that could guide cells collectively to use environmental cues, computational capacity, communication nodes to reinstate polarised state of cells. If we stick to this bottom-up approach, we may not be able to analyse the computational capacity & deliver instructions to the cell collective that could rebuild tissues or repair cells such as cancer cells. Even the genomic editing approaches will not work as it wont know which genes to edit to rebuild tissues while genes does not drive anatomical structure, they only provide instructions for which proteins have to be encoded for
The strategy of micromanaging cells- activating or inhibiting protein or triggering production of molecules is going to lead to “Genomic Winter”
While proponents of genome & DNA would not accept this as they have have never imagined biology beyond cellular & sub-cellular activities.Hardwired genome cannot drive predictions on the anatomical structures as they address sub-cellular level questions but does not decipher how cell collective communicate using the bioelectric network & works together towards large scale morphogenetic goals or re-establishing polarized state(resting membrane potential of cells).
These anatomical structures could drive the cognitive tasks of cell collectives that could be executed using informational processes embodying those tasks.
Take example of stem cells- stem cells once activated could be mobilised & differentiated into different cells to repair or replace dysfunctional cells or tissues. But the stem cell(s) do not work in isolation, nor do they work on single master commands. The stem cell differentiation works on the principle of collective intelligence. Individual stem cells sense the environmental cues & communicate with the cell collective. They use multi-scale competency architectures to process local mechanical and chemical cues. They detect growth factors, and electrical gradients, making split-second decisions to assume other cells. Cells communicate continuously through direct contact and the secretion of signaling molecules like proteins. The complex tissue pattern emerges from these simple, local interactions. If the development processes are disrupted, stem cells use chemical, physical, and mechanical signals to decide which specialized cell types they must become to interpret their location and differentiate into the appropriate tissue layer based on where they end up(2).
Yet there is a fundamental question here ?
When evolution lives on selfish motives, with each cell having its own self, how do these cells join & communicate through a bioelectric network & work towards large scale anatomical goals to maintain a morphogenetic structure & keep the host( our body) healthy? How does cooperation arise from the actions of these selfish agents?
The answer to these questions in prisoner’s dilemma(PD) – the game theory
Consider each grid of cells leveraging PD with its neighbouring cells. Let’s assume there are a fixed number of agentic cells & each one is distinct. Each one decides to cooperate or defect with its neighbours. A specific protein- gap junction allows cells to communicate with each other via small tunnels through which molecules can travel. This evolutionary discovery allows communication & cooperation within the cell collective. In this way, when two cells connect, they share nutrients & chemical & information signals via gap junction proteins. Since the cells are tied together, malpractices by one cell surely impact the other connected cell. This cooperation ensures that both cells work together for betterment of the host’s health. Every cell(agent) is as selfish as before but the difference is now there is a large self. This bioelectric network erases the boundaries between the cells & enhances the computational capacity of a group of cells-forming a supercell group where individual self of smaller subunits is hard to maintain. This has a massive impact from a game theory perspective with information processing & sharing through the entire cellular network. The incentive is clear-each cell gain from the learning, patterned memories & history of neighboring cells for which they have already paid a metabolic cost.
When cells link their bioelectric and signaling networks, something remarkable emerges — the computational processes that detect deviation from a target state become amplified across the collective. In large multicellular assemblies, the modest individual goals of single cells scale into coordinated tissue-level objectives, capable of driving regeneration toward a desired morphological state from virtually any starting configuration, correcting errors throughout the process.
Crucially, this collective decision-making is orchestrated through bioelectric communication networks — not the genome. While cells may draw on their genetic blueprints, accumulated learning, and bioelectrically encoded pattern memories, they are equally capable of overriding genetic instructions and compensating for genetic mutations — relying instead on intercellular communication to reconstruct the intended architecture of cells, tissues, and organs.
This cooperation is less about genetic /genome & rather about physiological interoperability. As long as Genomic hardware is good enough to drive the cellular communication, computational & information processing methodology of cellular collective. This means that if the human anatomical structure building software is intact, it can be used to build a right anatomical structure even if it is run on a completely different hardware(genome). The robust software- the cellular communication network will be instructed to build same structure even overriding the genetic instructions /overcoming mutations

At the cellular level, most cells are capable of forming networks with one another. This connectivity is governed by gap junctions — specialized proteins that act as channels between cells. Whether these channels are open or closed, and what molecules or signals pass through them, constitutes the cell’s functional “software” or computational logic. Crucially, gap junctions are not static; they carry a form of molecular memory, shaped by the prior states of the cells they connect. It is this memory that determines whether connexin proteins remain open and permit the exchange of molecules.
Gap junctions function as voltage-gated conductors operating through a memory-driven feedback loop, enabling cohesive, collective decision-making. Cells join larger cellular communities by sharing nutrients, expelling toxins, and exchanging information — all while employing predictive computation to sustain cellular balance and adaptability. Remarkably, cells can also collaborate to equalize voltage across the network through gap junction signaling. This process helps reintegrate aberrant cells — those in a depolarized state — back into the broader cell collective, redirecting them away from autonomous, potentially pre-symptomatic behavior.
Every single-celled organism possesses a form of basal cognition and a rudimentary mind. This allows it to leverage its limited local computational capacity to pursue simple, immediate goals — while collective organization dramatically amplifies that capacity, enabling far more complex goals essential to our health and wellbeing.
This principle — where cellular memory and predictive processing drive large-scale morphogenetic outcomes — is not confined to neurons. It applies equally to non-neuronal cells, suggesting that biological intelligence is far more distributed and fundamental than traditionally recognized.
To sum up, the focus of development biology combined with frontier technologies is not to focus on the blueprint instruction manual but how cells sense the environment, read past memories & genetic instructions to send signals, communicate bioelectrically & even override genetic instructions/overcome mutations to pursue grandiose human health goals. This has huge implications for cancer & would bring a paradigm shift in how we treat cancer- the future of cancer may not lie in killing those with aberrant signals but altering the voltage gradient of cells to make them rejoin the cell collective & work in a cohesive way to keep the human host healthy.
Mitochondria- The organelle that is playing a central role in regulating the ion channel & gap junction proteins
Most of us remember the classic high school biology line: “Mitochondria are the powerhouses of the cell.” While it’s true that these ancient organelles fuel our bodies with ATP, modern science has revealed they do so much more. In fact, energy production is just one of thousands of functions they perform.
The story of mitochondria began about two billion years ago when a larger ancestral cell engulfed a smaller bacterium. Instead of digesting it, the host cell formed a life-changing symbiotic partnership. It offered shelter and oxygen; in return, the bacterium provided a highly efficient way to generate energy using that oxygen, completely outperforming the slow fermentation models of the time. This alliance gave rise to eukaryotic cells, paving the way for all complex life—every plant, animal, and human can trace their lineage back to this single event.
Today, these ancient organelles live in almost every cell of our body (except red blood cells). With roughly 37 trillion cells hosting between 1,000 and 2,000 mitochondria each, your body houses an astonishing 7 to 14 quadrillion of them. That is a population far larger than all the bacteria in your gut and mouth combined.
For decades, we believed their only job was converting biochemical energy into electrochemical ATP to fund daily cellular tasks—like synthesizing proteins, activating enzymes, or producing hormones like vitamin D, insulin, and cortisol. However, groundbreaking research is flipping this paradigm on its head. We now know that mitochondria aren’t just factory workers; they are the ultimate controllers of our cells, even dictating instructions to our genome.
Operating as a massive, distributed bioelectric and information-processing network, these 14 quadrillion organelles use specialized receptors to sense the external environment. They integrate this data and release signaling molecules (like cytokines and mitokines) to communicate with other cells, driving physical adaptation. As active signaling hubs, they constantly communicate with nucleus about nutrient levels and oxidative stress. The nucleus then uses this feedback to alter gene expression and regulate our overall health.
Remarkably, mitochondria even have their own DNA. When they experience severe stress, they can leak this genetic material, which can integrate into our nuclear DNA—a cellular distress signal that often marks the onset of disease.
The energetic cost of stress(3)
The fundamental law of thermodynamics dictates that energy cannot be created or destroyed—only transformed. This principle is the bedrock of human biology. We operate on a strict, finite energy budget that must be carefully divided between two competing priorities:
- Vital Processes: Energy required for sustaining life
- (GMR): The essential energy allocated to Growth, Maintenance, and Repair.
- Stress Responses: The energy required to adapt to immediate threats.
The Cellular Alarm: Mitochondrial Communication
When the body encounters stressors—whether environmental toxins, nutritional deficiencies, or signals from a disrupted microbiome—our mitochondria act as the primary sensors. They hold cellular “roundtable conferences,” communicating dynamically across their network. To survive the threat, the body initiates an energy reallocation process, pulling precious resources away from expensive GMR tasks to fuel the immediate allostatic (adaptive) response.
The Shift: Chronic Stress and Allostatic Load
When these toxins and stressors become a constant presence, the body is forced to establish a new, dysfunctional baseline.This persistent activation of stress responses drastically increases daily energetic costs.
To cope with this relentless demand, the body undergoes cellular and physiological recalibrations. We begin to see early warning signs like hyperglycemia (elevated blood sugar) and elevated blood lipids—a state known as allostatic load.
The Breakdown: Allostatic Overload and Disease
Over time, sustaining this high-alert state alters our biology permanently. The chronic strain causes structural and functional damage to the body’s bioelectric and regulatory networks, manifesting as:
- High blood pressure
- Downregulation of hormone receptors
- Pathological remodeling of brain circuitry & more
Ultimately, these long-term adaptations cause a systemic breakdown in internal communication and regulation. This final stage—allostatic overload—directly translates to accumulated cellular damage, accelerated aging, and the onset of chronic disease.
Energy resistance principle
The energetic cost of stress offers a compelling lens through which to understand the principle of energy resistance within the mitochondrial electron transport chain. A certain degree of resistance is inherent to the process of energy conversion as electrons traverse the complexes of the electron transport chain (Complexes I–V).
However, when resistance is amplified by stressors or toxins, the system becomes dysregulated — electrons flow backward and electrons leak onto molecular oxygen, generating reactive oxygen species (ROS). The result is a dual burden: diminished ATP production efficiency and excess dissipative heat that agitates surrounding molecules, perpetuating further heat generation and ultimately inflicting DNA damage. The mutations that emerge under these conditions are a direct consequence of this heightened energetic resistance.
When mitochondria are exposed to stressors or toxins that elevate their stress cost, they do not respond in isolation. They communicate with neighboring mitochondria and cells through the release of signaling molecules — including cytokines such as GDF-15 and TNF-alpha — triggering a coordinated reallocation of energy toward managing those stressors. Sustained over time, this process accumulates as allostatic load, driving physiological recalibration and, eventually, disease onset.
Mitochondria also relay distress signals directly to the nucleus, instructing it to alter gene expression in ways that activate proteins and metabolites fueling inflammatory cascades — another pathway through which chronic stress translates into pathology.
In broader terms, when mitochondria detect metabolic shifts or stress, they dispatch molecular signals — calcium ions, ROS, or AMPK activation — that reach the nucleus and retroactively reshape the cell’s transcriptional programs.
The cell collective can then read these gene expression changes, or draw on its patterned cellular memory, to coordinate cell-to-cell communication and make decisions aligned with its anatomical objectives. These decisions govern whether cells differentiate, proliferate, or migrate into specific structural positions within a tissue.
It is precisely this dynamic interplay between mitochondrial signaling and collective gene expression that drives the structural and anatomical goals of the organism — enabling tissues to develop, sustain homeostasis, and repair themselves.
As discussed above, mitochondria acts as information processing & signal transduction network & instructs nucleus to alter gene expression These functions yield proteins such as connexins that build gap junctions & ion channels. This creates a direct feedback loop where mitochondria functions, metabolism & signalling how cell collective communicate electrically & chemically
It is important to remember that genetic mutations or altered gene expression are downstream of mitochondria functions & signals transduced by them.
Final words
The human body stands as nature’s most extraordinary and intelligent system — a living architecture built from remarkably capable fundamental units: cells. Each cell operates within a defined computational boundary, pursuing its own local objectives. Yet within the cell’s cytoplasm, mitochondria — ancient organelles responsible for energy transformation — perform functions that extend far beyond metabolism. They regulate the production of gap junction proteins and modulate ion channel activity, collectively maintaining the transmembrane potential that keeps cells electrically balanced.
Beyond the individual cell lies something far more powerful: the cell collective. This expansive cellular network operates at a vastly greater cognitive scale, possessing an amplified computational capacity that emerges through cooperation. Cells within this collective communicate through ion channels and gap junction proteins, allowing them to reference stored patterned memories and genetic instructions housed in the nucleus, while also responding to adaptive shifts in gene expression. Together, they coordinate anatomical goals and govern decisions surrounding cell growth, differentiation, division, and repair. Crucially, when genetic mutations or errors arise, the cell collective can orchestrate corrective responses — drawing on mitochondria-regulated gene expression to restore healthy physiological function.
External stressors and mitochondria-derived cytokines can destabilize ion channel equilibrium, pushing cells toward a depolarized state — one where the internal resting potential shifts from negative to positive. This disruption causes ion channels to cause electrostatic barriers and forces gap junctions to close, interfering with the cell collective’s communication network. The consequences can be severe: isolated cells, severed from collective signaling, may revert to primitive, pre-symbiotic behavioral patterns — pursuing self-serving, unilateral goals rather than contributing to the greater cellular community. This breakdown is increasingly understood as a foundational mechanism in the onset of cancer.
Shifting scientific focus toward developmental biology and the diverse intelligence embedded within cells — recognizing the genome as just one component of a broader cellular communication network rather than its sole master — could trigger a profound paradigm shift in oncology, and meaningfully improve cancer survival rates.
The most promising frontier in medicine may lie in moving beyond the constraints of classical molecular biology dogma, and instead directing innovation toward the bioelectric communication networks that silently govern the body’s most fundamental decisions.
Citations
https://www.cell.com/trends/molecular-medicine/fulltext/S1471-4914(23)00142-9
https://www.frontiersin.org/journals/psychology/articles/10.3389/fpsyg.2019.02688/full
https://pmc.ncbi.nlm.nih.gov/articles/PMC10082134/
https://pmc.ncbi.nlm.nih.gov/articles/PMC10082134/
https://pmc.ncbi.nlm.nih.gov/articles/PMC5901651
https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2023.1238171/full
https://aeon.co/essays/how-understanding-bioenergetics-can-help-our-brain-health
https://aeon.co/essays/how-to-understand-cells-tissues-and-organisms-as-agents-with-agendas
Leave a Reply